発生ゲノム科学研究室
私達の研究室(幸田研)は2018年度に発足した新しい研究室(教授:幸田 尚、助教:志浦 寛相)で、哺乳類の初期発生における遺伝子発現制御、胎盤形成機構の解明など、ヒトを含めた哺乳類の初期胚発生に重要な過程を明らかにすることを目指し、分子生物学的なアプローチを用いて研究を進めています。
①次世代シークエンサーを駆使した発生初期胚の遺伝子発現解析
不妊治療(生殖補助医療)を経て生まれる子供の数は年々増え続けており、現在では毎年3万人(35人に一人)の子供が体外受精によって生まれています。不妊の原因となりうる事象としては、親年齢の高齢化、親の偏った栄養・食生活などが考えられます。私たちはそのような状況が発生初期の胚にどのような影響を与えるのかを「次世代シーケンスによる全遺伝子発現解析」により調べることで、不妊の原因となる遺伝子発現異常を特定し、生殖補助医療の向上へとつなげることを目標としています。

②ヒトを含む哺乳類はどのように生まれたか?−哺乳類の進化(哺乳類らしさ)に関わる遺伝子の研究−
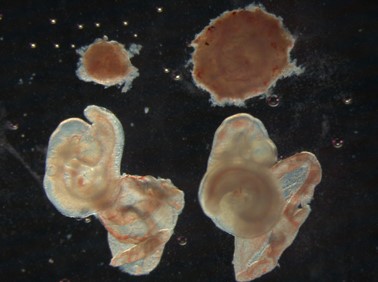
これまでの研究から哺乳類の誕生と同じ頃にゲノム中に新しく獲得されたと考えられるPeg10という遺伝子が胎盤を作るのに必須な遺伝子であることが分かりました(写真:右:正常マウス、左:Peg10遺伝子を欠損させたマウス。Peg10欠損マウスでは写真上部にある胎盤(丸い組織)が未発達であることが分かる)。胎盤は哺乳類だけが持ち、お腹の中で子どもを育てる「胎生」という発生様式に不可欠な組織です。私たちはこの新しいPeg10という遺伝子の機能解析を通じて、胎盤組織の発生・形成メカニズムの解明と同時に、この遺伝子の獲得が哺乳類独自の胎生という発生様式の進化にどのように関わったのかを明らかにしたいと考えています。
③ A・C・G・Tだけではない第5・6の塩基を解析する新技術の開発
ゲノムDNAを構成する塩基はA・C・G・Tの4種類あり、この並び方により遺伝情報が記述されています。しかし、私たちのゲノムの中にはこれ以外に、「C」の一部が修飾された「メチルシトシン(=第5の塩基)」と「ヒドロキシメチルシトシン(=第6の塩基)」が存在し、これらが遺伝子発現のON/OFFを制御していることが分かってきました。しかし、これまでのDNA塩基解析法では、普通の「C」と第5・6の塩基である「メチルシトシン(mC)」「ヒドロキシメチルシトシン(hmC)」を区別することができませんでした。私たちは、この「メチルシトシン」「ヒドロキシメチルシトシン」を含む6種類の塩基の全てを区別する新規の塩基解析技術を開発しています(図:EnIGMA法)。
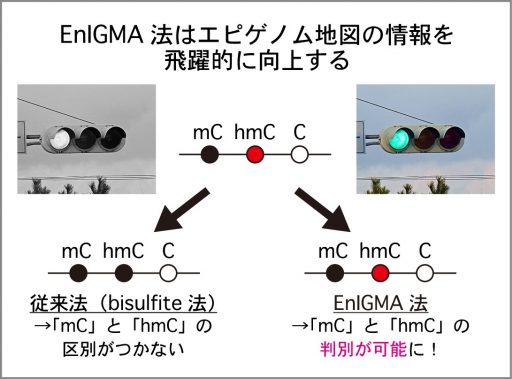
これはちょうど白黒写真がカラー写真になると大きく情報量が増えるのと同じで、遺伝子発現のON/OFFこれまで以上にはっきりと解析できるようになる可能性のある画期的な技術です。
幸田研は若山研・岸上研と共に発生工学研究センターに所属しており、最新の設備を用いた研究が可能なだけでなく、研究室同士の連携のもと、多くの教員・学生とディスカッションできる環境が整っています。
意欲のある学生さんを大募集しています。ぜひ私たちといっしょに研究しましょう!